
Gli amminoacidi, e non le proteine in sé, sono i nutrienti necessari. Gli amminoacidi assorbiti ed utilizzati principalmente come elementi costitutivi per la sintesi delle proteine sono vitali per il mantenimento, la crescita, la riproduzione e la produzione di latte.
Nutrizione proteica
La nutrizione proteica nei ruminanti si è evoluta negli ultimi decenni a partire dalla determinazione del fabbisogno di proteine grezze (CP) negli anni ’70, quando la ricerca si è concentrata su quante proteine fossero necessarie per il rumine e quante per la manutenzione generale e la produzione di latte nella bovina. Nel corso del tempo, una migliore comprensione del metabolismo delle proteine ha permesso di calcolare il fabbisogno specifico di proteine nella dieta (N), che può essere classificato in due grandi categorie (NRC, 2001):
- Rumen Degradable Protein (RDP) o Proteina Degradabile Ruminale. Si tratta di proteine alimentari degradate dai microrganismi del rumine in presenza di un adeguato apporto energetico che forniscono la maggior parte dell’Azoto (N) necessario per la sintesi di Microbial Crude Protein (MCP) o Proteina Microbica Grezza.
- Rumen Undegradable Protein (RUP) o Proteina rumino-indegradabile. Si tratta della proteina alimentare che sfugge alla degradazione nel rumine e viene digerita dagli enzimi dell’animale ospite nell’intestino tenue. Oltre alla vera proteina, la RDP include anche l’azoto non proteico (NPN).
 La microflora ruminale passa continuamente dal rumine all’abomaso e all’intestino tenue dove muore e viene digerita dagli enzimi presenti in questo tratto di intestino per fornire i singoli aminoacidi (AA). Gli aminoacidi derivano anche dalla digestione della frazione RUP. I ruminanti assorbono poi i singoli aminoacidi. Quando queste due fonti di aminoacidi sono digerite e assorbite nell’intestino tenue vengono indicate collettivamente come Metabolizable Protein (MP), o Proteine Metabolizzabili.
La microflora ruminale passa continuamente dal rumine all’abomaso e all’intestino tenue dove muore e viene digerita dagli enzimi presenti in questo tratto di intestino per fornire i singoli aminoacidi (AA). Gli aminoacidi derivano anche dalla digestione della frazione RUP. I ruminanti assorbono poi i singoli aminoacidi. Quando queste due fonti di aminoacidi sono digerite e assorbite nell’intestino tenue vengono indicate collettivamente come Metabolizable Protein (MP), o Proteine Metabolizzabili.
Ruolo degli amminoacidi
È noto da tempo che le vacche hanno bisogno di aminoacidi per la sintesi del proprio tessuto corporeo e delle proteine regolatorie, protettive e secretorie: migliaia di proteine diverse devono essere sintetizzate ogni giorno. È anche ben documentato che la composizione in aminoacidi di ogni proteina è diversa, ma la composizione in aminoacidi di una proteina è la stessa ogni volta che viene sintetizzata perché la sintesi proteica è un evento geneticamente determinato. Pertanto, la composizione in aminoacidi delle proteine sintetizzate non è influenzata dalla quantità o dal profilo aminoacidico degli aminoacidi assorbiti.
Gli aminoacidi, e non le proteine in sé, sono i nutrienti necessari per i ruminanti. Gli aminoacidi assorbiti, utilizzati principalmente come elementi costitutivi per la sintesi delle proteine, sono vitali per il mantenimento, la crescita, la riproduzione e l’allattamento dei bovini da latte (NRC, 2001). Gli aminoacidi sono essenziali in quanto rappresentano i mattoni necessari per la sintesi dei tessuti e delle proteine del latte. Inoltre, sebbene in misura minore, gli aminoacidi assorbiti servono anche come precursori per la sintesi di altri metaboliti. Gli aminoacidi sono anche precursori della gluconeogenesi e possono essere convertiti in acidi grassi o servire come fonti immediate di energia metabolica quando vengono ossidati.

Equilibrio aminoacidico
Mentre il profilo degli aminoacidi assorbiti non influenza gli aminoacidi utilizzati nel metabolismo e nella produzione, quello degli aminoacidi essenziali (EAA) assorbiti influenza la quantità di proteine che possono essere sintetizzate. Questo fenomeno è ampiamente dimostrato nei suini (NRC, 1998) e nel pollame (NRC, 1994). È ben documentato che, quando gli EAA vengono assorbiti come richiesto dall’animale, la loro efficienza di utilizzo per la sintesi proteica è massimizzata e la richiesta di aminoacidi totali assorbiti è ridotta al minimo. Così, il catabolismo degli aminoacidi “rimasti” è minimo e l’escrezione di N urinaria è ridotta. Al contrario, l’efficienza di utilizzo degli aminoacidi per il metabolismo e la produzione delle proteine è meno efficiente quando il profilo di EAA assorbiti non soddisfa i fabbisogni ideali. In questo caso, sarà l’apporto del primo EAA limitante a determinare l’entità della sintesi proteica, non l’apporto totale di aminoacidi.
Aminoacidi limitanti
Gli aminoacidi “limitanti” sono quelli che scarseggiano rispetto alla quantità necessaria: nello specifico si tratta di Lisina (Lys), Metionina (Met) e Istidina (His). Questi AA sono stati riconosciuti a livello globale come i primi tre aminoacidi limitanti nella nutrizione delle vacche da latte. In particolare, Lys e Met sono gli amminoacidi più frequentemente identificati come i due più limitanti per la lattazione. Una varietà di studi di ricerca continua a sostenere queste prime osservazioni (ad esempio, Appuhamy et al., 2011; Chen et al., 2011; Doepel e Lapierre, 2010, 2011; Noftsger e St-Pierre, 2003; Noftsger et al., 2005; Ordway et al., 2009; Socha et al., 2005; St-Pierre e Sylvester, 2005). La Met è in genere il primo aminoacido limitante quando la maggior parte della RUP è apportata dalle proteine della soia (NRC, 2001). Queste fonti proteiche (Tabella 1) hanno basse concentrazioni di Met rispetto al latte e alle proteine batteriche. Come mostrato nella Tabella 1, la Met nel latte e nei batteri è rispettivamente pari al 2,6 e 2,7% del valore della proteina grezza (CP), mentre nella soia, nel sangue e nelle farine di piume e di carne la Met è sempre compresa in un range che varia dallo 0,8 all’1,4% del valore della CP.
La Lys è in primo luogo “limitante essenziale” quando il mais e altri cereali, e loro derivati, forniscono la maggior parte o la totalità della RUP (NRC, 2001). La lisina nel latte e nei batteri è rispettivamente il 7,6 e il 7,9% della CP, mentre nell’insilato di mais, nel mais, nei distillers di mais e nella farina di glutine di mais, il valore in Lys è sempre compreso in un range che varia dall’1,7 al 2,8% della CP.
La Lys è identificata come il primo AA “limitante” quando l’insilato di erba, l’ orzo e l’avena sono impiegate nella dieta in apporti considerevoli (Kim et al., 1999, 2000, 2001a, 2001b; Huhtanen et al., 2002; Korhonen et al., 2000).
Stima degli EAA nella MP
Le stime iniziali delle concentrazioni ideali di EAA espresse sulla Proteina Metabolizzabile (MP) sono state stabilite da Rulquin et al., (1993), NRC (2001) e Doepel et al. (2004). I diversi approcci hanno fornito raccomandazioni sorprendentemente simili per le forniture ottimali di Lys e Met in relazione alla MP. Recentemente, Schwab et al. (2009) hanno ripetuto i diagrammi dose-risposta di Lys e Met utilizzando la versione finale del modello NRC (2001), piuttosto che la versione beta utilizzata in precedenza. Sono stati utilizzati gli stessi studi e sono stati ripetuti tutti i passaggi indicati nei quali i prodotti derivati da orzo e frumento sostituiscono quantità significative di mais nella dieta in NRC (2001). In modo simile, Whitehouse et al. (2009) hanno ripetuto gli stessi passi, utilizzando gli stessi studi utilizzati per NRC (2001), per generare diagrammi dose-risposta di Lys e Met per i modelli matematici dinamici di CPM-Dairy e AMTS (software di formulazione della razione di ruminanti). Questo è stato fatto per entrambi i modelli basati sul CNCPS (Net Carbohydrate e Protein System), a causa della loro ampia diffusione nell’industria lattiero-casearia e per la preoccupazione che gli utenti di questi modelli possano utilizzare in modo non corretto le raccomandazioni generate utilizzando il modello NRC (Tabella 3).
Tabella 1. Un confronto della composizione aminoacidica essenziale dei tessuti magri del corpo, del latte e dei batteri ruminali con quella di alcuni alimenti zootecnici comuni1.


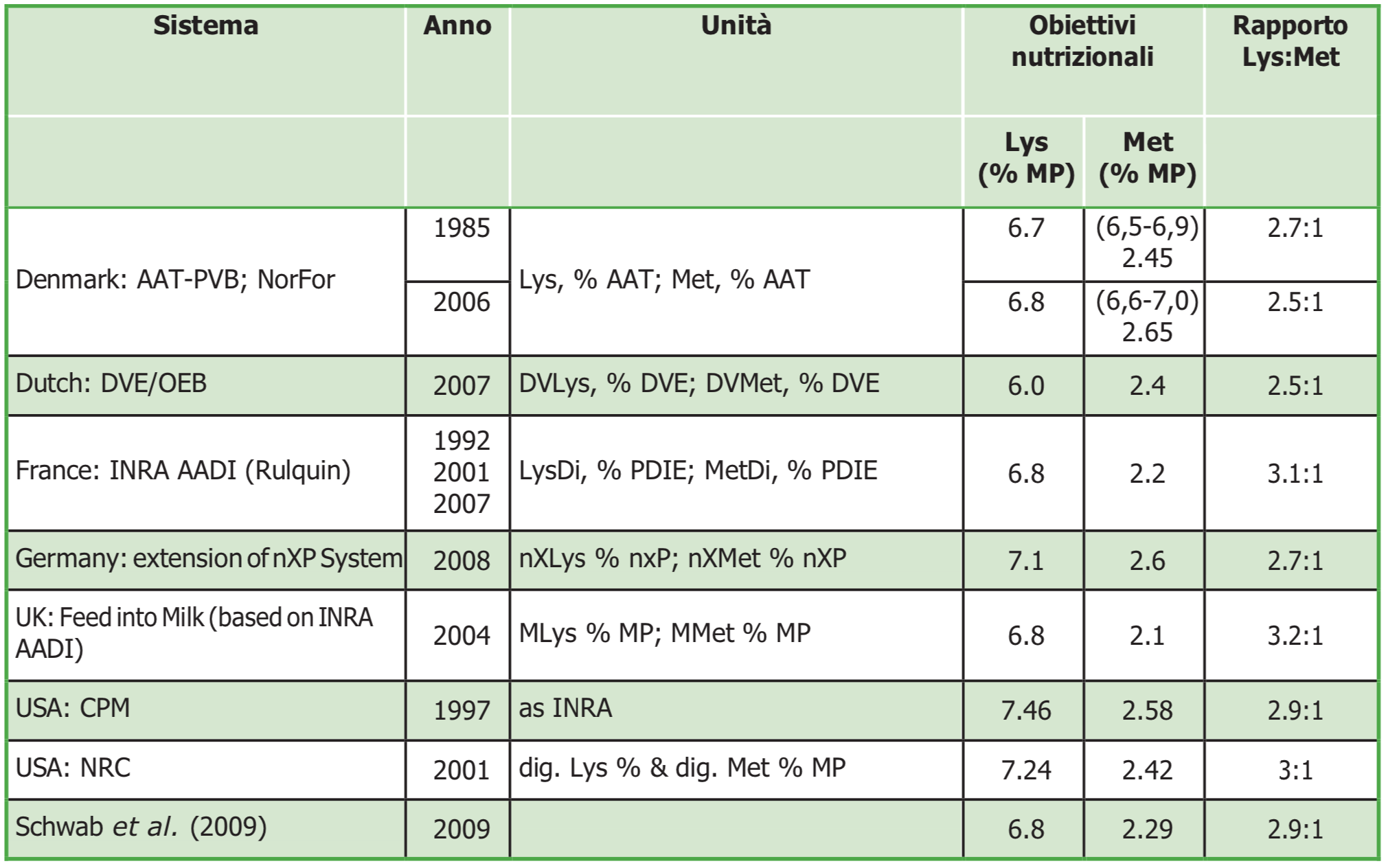
Tabella 2. Concentrazione ideale del rapporto tra Lys e Met espressa in % sulla MP per la produzione di proteine del latte in bovine in lattazione in diversi modelli di razionamento.
Analisi delle strategie alimentari proteiche in relazione al bilanciamento degli amminoacidi nella razione
– Massimizzare la sintesi di proteine microbiche (MCP) (ma non sovralimentando mediante un aumento in RDP nella dieta). Come mostrato nella Tabella 1, le proteine microbiche contengono alte percentuali di lisina e metionina. Aumentando la quantità di proteina microbica si riduce il divario tra gli EAA richiesti e quelli forniti.
– Non sovralimentare per la frazione RUP. L’alimentazione eccessiva in RUP ha diversi svantaggi nelle vacche da latte. La RUP in eccesso, di solito, sostituisce i carboidrati fermentescibili nella dieta come substrati primari per la sintesi dei componenti del latte. Essa riduce il contenuto di RDP al di sotto del livello raccomandato, e porta ad ottenere delle concentrazioni più basse di Lys e Met (% MP) e comporta spesso una dieta più costosa.
– Fornire una fonte di RUP bilanciata per Lys e Met. Una volta fissato il profilo aminoacidico, l’altra porzione di MP è costituita dalla RUP. Tuttavia, la RUP degli alimenti zootecnici è generalmente bassa in Lys, Met o entrambi. Gli aminoacidi rumino-protetti sono una soluzione efficiente per ottimizzare il rapporto e il livello di Lys e Met (% MP).
– Mantenere il rapporto di lisina e metionina. Per il massimo utilizzo degli aminoacidi assorbiti per la produzione e le funzioni metaboliche, il rapporto Lys:Met deve essere mantenuto secondo il sistema proteico di riferimento adottato (Tabella 2).
I benefici del bilanciamento aminoacidico… prossimamente
Le seguenti strategie e benefici verranno discusso in dettaglio nei prossimi capitoli:
- Aumentare la produzione di latte e dei componenti del latte.
- Migliorare l’efficienza dell’utilizzo dell’azoto nei mangimi.
- Migliorare le prestazioni riproduttive e la salute del ruminante.
- Ridurre l’escrezione di N per unità di latte, o di proteine del latte, prodotte.
- Aumentare la redditività del bestiame da latte.
Autori
Mohammad Al Rifai, Stefano Mattuzzi
Riferimenti
Appuhamy, J. A. D. R. N., J. R. Knapp, O. Becvar, J. Escobar, and D. Hanigan. 2011. Effects of jugular-infused lysine, methionine, and branched-chain amino acids on milk protein synthesis in high-producing dairy cows. J. Dairy Sci. 94: 1952–1960.
Chen, Z. H., G. A. Broderick, N. D. Luchini, B. K. Sloan, and Devillard. 2011. Effect of feeding different sources of rumen-protected methionine on milk production and N-utilization inlactating dairy cows. J. Dairy Sci. 94: 1978–1988.
Doepel, L., D. Pacheco, J. J. Kennelly, M. D. Hanigan, I. F. Lopez, and H. Lapierre. 2004. Milk protein synthesis as a function of amino acid supply. J. Dairy Sci. 87: 1279–1297.
Doepel, L. and H. Lapierre. 2010. Changes in production and mammary metabolism of dairy cows in response to essential and nonessential amino acid infusions. J. Dairy Sci. 93: 3264–3274.
Doepel, L. and H. Lapierre. 2011. Deletion of arginine from an abomasal infusion of amino acids does not decrease milk protein yield in Holstein cows. J. Dairy Sci. 94: 864-873.
Huhtanen, P., V. Vanhatalo, and T. Varvikko. 2002. Effects of abomasal infusions of histidine, glucose, and leucine on milk production and plasma metabolites of dairy cows fed grass silage diets. J. Dairy Sci. 85: 204-216.
Kim, C. H., T. G. Kim, J. J. Choung, and D. G. Chamberlain. 1999. Determination of the first limiting amino acid for milk production in dairy cows consuming a diet of grass silage and a cereal-based supplement containing feather meal. J. Sci. Food Agric. 79: 1703-1708.
Kim, C. H., T. G. Kim, J. J. Choung, and D. G. Chamberlain. 2000. Variability in the ranking of the three most-limiting amino acids for milk protein production in dairy cows consuming grass silage and a cereal-based supplement containing feather meal. J. Sci. Food Agric. 80: 1386-1392
Kim, C. H., T. G. Kim, J. J. Choung, and D. G. Chamberlain. 2001a. Effects of intravenous infusion of amino acids and glucose on theyield and concentration of milk protein in dairy cows. J. Dairy Res. 68: 27-34.
Kim, C. H., T. G. Kim, J. J. Choung, and D. G. Chamberlain. 2001b. Estimates of the efficiency of transfer of L-histidine from bloodto milk when it is the first-limiting amino acid for secretion of milk protein in the dairy cow. J. Sci. Food Agric. 81: 1150-1155.
Korhonen, M., A. Vanhatalo, T. Varvikko, and P. Huhtanen. 2000. Responses to graded doses of histidine in dairy cows fed grass silage diets. J. Dairy Sci. 83: 2596-2608.
McLaughlin, A. M. 2002. Evaluation of lysine supplements for lactating cows. M.S. Thesis, University of New Hampshire. 159 pp.
National Research Council. 1994. Nutrient Requirements of Poultry. 9th rev. ed. Natl. Acad. Sci., Washington, DC.
National Research Council. 1998. Nutrient Requirements of Swine. 10th rev. ed. Natl. Acad. Sci., Washington, DC.
National Research Council. 2001. Nutrient Requirements of Dairy Cattle, 7th rev. ed. Washington, D.C.: National AcademyPress.
Noftsger, S. and N. R. St-Pierre. 2003. Supplementation of methionine and selection of highly digestible rumenundegradable protein to improve nitrogen efficiency for milk production. J. Dairy Sci. 86: 958-969.
Noftsger, S., N. R. St-Pierre and J. T. Sylvester. 2005. Determination of rumen degradability and ruminal effects of threesources of methionine in lactating cows. J. Dairy Sci. 88: 223–237.
O’Connor, J. D., C. J. Sniffen, D. G. Fox, and W. Chalupa. 1993. A net carbohydrate and protein system for evaluating cattle diets: Predicting amino acid adequacy. J. Anim. Sci. 71: 1298-1311.
Ordway R. S., S. E. Boucher, N. L. Whitehouse, C. G. Schwab, and K. Sloan. 2009. Effects of providing two forms of supplemental methionine to periparturient Holstein dairy cows on feed intakeand lactational performance. J. Dairy Sci. 92: 5154-5166.
Rulquin, H., P. M. Pisulewski, R. Verite, and J. Guinard. 1993. Milk production and composition as a function of postruminal lysine and methionine supply: A nutrient response approach. Livest. Prod. Sci. 37: 69-90.
Schwab, C., N. Whitehouse, D. Luchini, and B. Sloan. 2009. Reevaluation of the breakpoint estimates for the NRC (2001) required concentrations of lysine and methionine in metabolizable protein for maximal content and yield of milk protein. J. DairySci. 92 (Suppl. 1):103. (Abstr.).
Socha, M. T., D. E. Putnam, B. D. Garthwaite, N. L. Whitehouse, A. Kierstead, C. G. Schwab, G. A. Ducharme, and J. C. Robert. 2005. Improving intestinal amino acid supply of pre- and postpartum dairy cows with rumen-protected methionine and lysine. J. Dairy Sci. 88: 1113-1126.
St-Pierre, N. R., and J. T. Sylvester. 2005. Effects of 2-hydroxy-4-(methylthio) butanoic acid (HMB) and its isopropyl ester onmilk production and composition by Holstein cows. J. Dairy Sci. 88: 2487-2407.
Huhtanen, P., V. Vanhatalo, and T. Varvikko. 2002. Effects of abomasal infusions of histidine, glucose, and leucine on milk production and plasma metabolites of dairy cows fed grass silage diets. J. Dairy Sci. 85: 204-216.
Whitehouse, N., C. Schwab, D. Luchini, T. Tylutki, and B. Sloan. 2009. Comparison of optimal lysine and methionine concentrations inmetabolizable protein estimated by the NRC 2001, CPM-Dairy (v.3.0.10) and AMTS. Cattle (v.2.1.1) models. J. Dairy Sci. 92 (Suppl. 1):103.(Abstr.)




